 物理與化學性質
物理與化學性質
脫氧核糖核酸
脫氧核醣核酸
(英語:deoxyribonucleic acid,縮寫:DNA)又稱去氧核醣核酸, 是一種生物大分子,可組成遺傳指令,引導生物發育與生命機能運作。
主要功能是資訊儲存,可比喻為「藍圖」或「食譜」。
其中包含的指令,是建構細胞內其他的化合物,如蛋白質與核醣核酸所需。
帶有蛋白質編碼的DNA片段稱為基因。
其他的DNA序列,有些直接以本身構造發揮作用,有些則參與調控遺傳訊息的表現。
DNA是一種長鏈聚合物,組成單位稱為核苷酸,而糖類與磷酸藉由酯鍵相連,組成其長鏈骨架。
每個糖單位都與四種鹼基裡的其中一種相接,這些鹼基沿著DNA長鏈所排列而成的序列,
可組成遺傳密碼,是蛋白質胺基酸序列合成的依據。
讀取密碼的過程稱為轉錄,是根據DNA序列複製出一段稱為RNA的核酸分子。
多數RNA帶有合成蛋白質的訊息,另有一些本身就擁有特殊功能,例如核糖體RNA、小核RNA與小干擾RNA。
在細胞內,DNA能組織成染色體結構,整組染色體則統稱為基因組。
染色體在細胞分裂之前會先行複製,此過程稱為DNA複製。
對真核生物,如動物、植物及真菌而言,染色體是存放於細胞核內;
對於原核生物而言,如細菌,則是存放在細胞質中的擬核裡。
染色體上的染色質蛋白,如組織蛋白,能夠將DNA組織並壓縮,
以幫助DNA與其他蛋白質進行交互作用,進而調節基因的轉錄。
物理與化學性質
脫氧核醣核酸是一種由核苷酸重複排列組成的長鏈聚合物,
寬度約22到24埃(2.2到2.4奈米),
每一個核苷酸單位則大約長3.3埃(0.33奈米)。
在整個脫氧核醣核酸聚合物中,可能含有數百萬個相連的核苷酸。
例如人類細胞中最大的1號染色體中,就有2億2千萬個鹼基對。
通常在生物體內,脫氧核醣核酸並非單一分子,
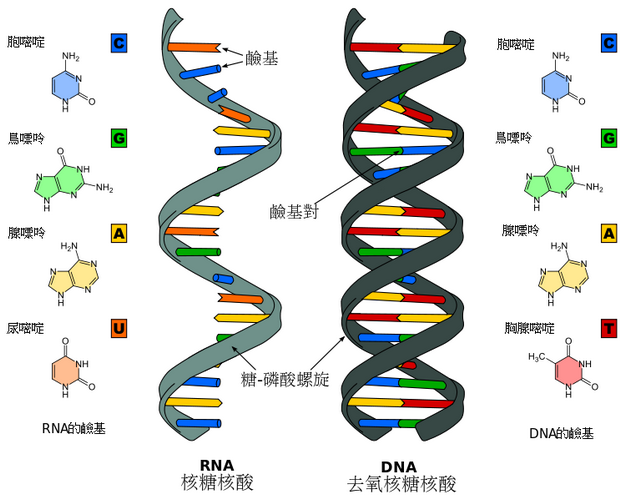
而是形成兩條互相配對並緊密結合,且如藤蔓般地纏繞成雙螺旋結構的分子。
每個核苷酸分子的其中一部分會相互連結,組成長鏈骨架;
另一部分稱為鹼基,可使成對的兩條脫氧核醣核酸相互結合。
所謂核苷酸,是指一個核苷加上一個或多個磷酸基團,
核苷則是指一個鹼基加上一個糖類分子。
脫氧核醣核酸骨架是由磷酸與糖類基團交互排列而成。
組成脫氧核醣核酸的糖類分子為環狀的2-去氧核醣,屬於五碳糖的一種。
磷酸基團上的兩個氧原子分別接在五碳糖的3號及5號碳原子上,
形成磷酸雙酯鍵。
這種兩側不對稱的共價鍵位置,使每一條脫氧核醣核酸長鏈皆具方向性。
雙螺旋中的兩股核苷酸互以相反方向排列,這種排列方式稱為反平行。
脫氧核醣核酸鏈上互不對稱的兩末端一邊叫做5'端,另一邊則稱3'端。
脫氧核醣核酸與RNA最主要的差異之一,在於組成糖分子的不同,DNA為2-去氧核醣,RNA則為核醣。
兩股脫氧核醣核酸長鏈上的鹼基以氫鍵相互吸引,使雙螺旋形態得以維持。
這些鹼基可分為兩大類,以5角及6角雜環化合物組合而成的一類稱為嘌呤;只有一個6角雜環的則稱為嘧啶。
組成脫氧核醣核酸的鹼基,分別是
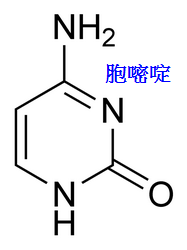
腺嘌呤(Adenine,縮寫A)、胞嘧啶(Cytosine,C)、 鳥嘌呤(Guanine,G)與胸腺嘧啶(Thymine,T)。
鹼基、糖類分子與磷酸三者結合之後,便成為完整的核苷酸。
還有一種鹼基稱為尿嘧啶(Uracil,U),此種鹼基比胸腺嘧啶少了一個位於環上的甲基, 一般出現在RNA分子中,
角色相當於脫氧核醣核酸裡的胸腺嘧啶。
通常在脫氧核醣核酸中,它會作為胞嘧啶的分解產物,或是CpG島中未經甲基化的胞嘧啶突變產物。
鹼基配對
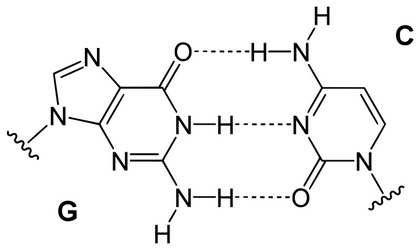
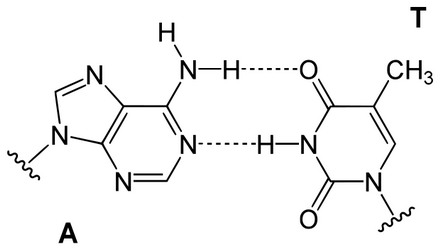
一股脫氧核醣核酸上所具有的各類型含氮鹼基,都只會與另一股上的一個特定類型鹼基產生鍵結。
此種情形稱為互補性鹼基配對。嘌呤與嘧啶之間會形成氫鍵,在一般情況下,A只與T相連,而C只與G相連。
因此排列於雙螺旋上的核苷酸,便以這種稱為鹼基對的方式相互聯結。
除此之外,與脫氧核醣核酸序列無關的疏水性效應,以及π重疊效應所產生的力,也是兩股脫氧核醣核酸能維持結合狀態的原因。
由於氫鍵比共價鍵更容易斷裂,這使雙股脫氧核醣核酸可能會因為機械力或高溫作用,而有如拉鍊一般地解開,這種現象被稱為DNA變性。
由於互補的特性,使位於雙股序列上的訊息,皆以雙倍的形式存在,這種特性對於脫氧核醣核酸複製過程來說相當重要。
互補鹼基之間可逆且具專一性的交互作用,是生物脫氧核醣核酸所共同擁有的關鍵功能。
兩種不同的鹼基對分別是以不同數目的氫鍵結合:A-T之間有兩條;G-C之間則有三條。
多一條氫鍵使GC配對的穩定性高於AT配對,也因此兩股脫氧核醣核酸的結合強度,是由GC配對所佔比例,以及雙螺旋的總長度來決定。
當脫氧核醣核酸雙螺旋較長且GC含量較高時,其雙股之間的結合能力較強;長度較短且AT含量較高時,結合能力則較弱。
雙螺旋上有某些部位必須能夠輕易解開,這些部位通常含有有較多的AT配對,例如細菌啟動子上一段含有TATAAT序列的普裡布諾盒。
在實驗室中,若找出解開氫鍵所需的溫度,也就是所謂熔點(Tm),便能計算出兩股之間的結合強度。
當脫氧核醣核酸雙螺旋上所有的鹼基配對都解開之後,溶液中的兩股脫氧核醣核酸將分裂成獨立的分子。
單股脫氧核醣核酸分子並無固定的形體,但仍有某些形狀較為穩定且常見。
正意與反意
一般來說,當一段脫氧核醣核酸序列為合成信使RNA(mRNA,可轉譯成蛋白質)所需時,稱為「正意」。
而相對並互補的另一股序列,則稱為「反意」。
由於RNA聚合酶的作用方式,是根據模板上的訊息來合成一段與模板互補的RNA片段,
因此正意mRNA的序列實際上與脫氧核醣核酸上的反意股相同。
在同一股脫氧核醣核酸上,可能同時會有屬於正意和反意的片段。
此外,反意RNA在原核生物或真核生物體內皆存在,但是其功能尚未明瞭。
有研究認為,反意RNA可利用RNA與RNA之間的鹼基配對,來調控基因的表現。
少數屬於原核生物、真核生物、質體或病毒的脫氧核醣核酸序列(後兩者較前兩者多),
會由於正意股與反意股之間的差異難以區分,而產生重疊基因,這類脫氧核醣核酸序列具有雙重功能,
一方面能以5'往3'的方向合成蛋白質,另一方面也能以相反方向合成另一個蛋白質。
這種重疊現象一方面在細菌體內參與調控基因的轉錄,一方面則在較小的病毒基因組中,扮演增加訊息量的角色。
為了縮減基因組的大小,也有某些病毒以線狀或環狀的單股脫氧核醣核酸作為遺傳物質。
超螺旋
脫氧核醣核酸鏈在雙螺旋基礎上如繩索般扭轉的現象與過程稱為DNA超螺旋。
當脫氧核醣核酸處於「鬆弛」狀態時,雙螺旋的兩股通常會延著中軸,以每10.4個鹼基對旋轉一圈的方式扭轉。
但如果脫氧核醣核酸受到扭轉,其兩股的纏繞方式將變得更緊或更鬆。
當脫氧核醣核酸扭轉方向與雙股螺旋的旋轉方向相同時,稱為正超螺旋,此時鹼基將更加緊密地結合。
反之若扭轉方向與雙股螺旋相反,則稱為負超螺旋,鹼基之間的結合度會降低。
自然界中大多數的脫氧核醣核酸,會因為拓撲異構酶的作用,而形成輕微的負超螺旋狀態。
拓撲異構酶同時也在轉錄作用或DNA複製過程中,負責紓解脫氧核醣核酸鏈所受的扭轉壓力。
由左到右分別為A型、B型與Z型三種脫氧核醣核酸結構。
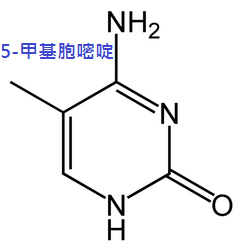
化學修飾
基因的表現,受染色體上的染色質結構與異染色質(基因無表現或低表現)區域裡的胞嘧啶甲基化所影響。
舉例而言,當胞嘧啶受到甲基化時,會轉變成5-甲基胞嘧啶,此作用對於X染色體的去活化、
銘印和保護脫氧核醣核酸分子不被內切酶所切斷(存在例外)而言相當重要。
甲基化的程度在不同生物之間有所差異,如秀麗隱桿線蟲便缺乏胞嘧啶甲基化,
而在脊椎動物體內則較常出現,大約有1%的脫氧核醣核酸為5-甲基胞嘧啶。
5-甲基胞嘧啶容易因自然發生的脫氨作用而變成胸腺嘧啶,也因此使甲基化的胞嘧啶成為突變熱點,
這也解釋了為什麼胞嘧啶和鳥嘌呤會集中出現在CpG島里,
因為那裡的甲基化作用被壓制,沒有甲基化的胞嘧啶所產生的突變產物並非胸腺嘧啶,而是尿嘧啶。
因為尿嘧啶會相對容易地被更正過來,所以CpG島內胞嘧啶不易形成突變而會被保留下來。
其他的鹼基修飾還包括細菌的腺嘌呤甲基化,以及使動質體(一種生物)的尿嘧啶轉變成「J-鹼基」的糖基化等。
苯並芘是一種突變原,可於菸葉燃燒生成的煙中發現,圖為苯並芘與脫氧核醣核酸的加合物。
脫氧核醣核酸損害
有許多不同種類的突變原可對DNA造成損害,其中包括氧化劑、烷化劑,以及高頻電磁輻射,如紫外線與X射線。
不同的突變原對DNA造成不同類型的損害,舉例而言,紫外線會造成胸腺嘧啶二聚體的形成,並與相鄰的鹼基產生交叉,進而使DNA發生損害。
另一方面,氧化劑如自由基或過氧化氫,可造成多種不同形態的損害,尤其可對鳥苷進行鹼基修飾,並且使雙股分解。
根據估計,在一個人類細胞中,每天大約有500個鹼基遭受氧化損害。
在各種氧化損害當中,以雙股分解最為危險,此種損害難以修復,且可造成DNA序列的點突變、插入與刪除,以及染色體易位。
許多突變原可嵌入相鄰的兩個鹼基對之間,這些嵌入劑大多是芳香性分子及平面分子,包括乙錠、道諾黴素、阿黴素與沙利竇邁。
必須先使鹼基之間的空隙變大,才能使嵌入劑置入鹼基對之間,整體而言,脫氧核醣核酸會因為雙螺旋解開而扭曲變形。
結構改變會使轉錄作用與脫氧核醣核酸複製過程受到抑制,進而導致毒害與突變。
因此脫氧核醣核酸嵌入劑通常也是致癌物,常見的例子有二醇環氧苯並芘、吖啶、黃麴毒素與溴化乙錠等。
但是這些物質也因為能夠抑制脫氧核醣核酸的轉錄與複製,而可應用於化學治療中,用以抑制癌症細胞的快速生長情形。
生物機能摡觀
脫氧核醣核酸於真核生物細胞內,通常是以長條狀染色體形式存在; 在原核生物細胞內則是環狀染色體。
細胞內的所有染色體合稱基因組。
人類基因組中大約有30億個鹼基對,共組成了46個染色體。
脫氧核醣核酸所攜帶的訊息,是以脫氧核醣核酸序列形式,保存於一些稱為基因的片段中。
基因中的遺傳訊息是經由互補的鹼基配對來傳遞,例如在轉錄作用中,
細胞裡的RNA核苷酸會與互補的脫氧核醣核酸結合,複製出一段與脫氧核醣核酸序列互補的RNA序列。
一般來說,這段RNA序列將會在轉譯作用中,經由RNA之間的互補配對,合成出相對應的蛋白質序列。
另一方面,細胞也可以在稱為脫氧核醣核酸複製的過程中,單純地複製其自身的遺傳訊息。
基因組結構
真核生物的基因組脫氧核醣核酸主要存放於細胞核中,此外也有少量位於粒線體或葉綠體內。
原核生物的脫氧核醣核酸則是保存在形狀不規則的類核(nucleoid)當中。
基因是脫氧核醣核酸的一段區域,保存了基因組裡的遺傳訊息,是遺傳的單位,影響了生物個體的特定表徵。
基因中含有可轉錄的開放閱讀框架,以及一些可調節開放閱讀框架表現的調控序列,如啟動子與強化子。
許多物種的基因組都只有一小部分可編譯成蛋白質。
以人類為例,在人類的基因組中只有1.5%屬於含有蛋白質編碼的外顯子,另有超過50%屬於無編碼的重複序列。
真核生物基因組中如此大量的非編碼DNA,以及物種之間不尋常的基因組大小或C值差異,
長久以來一直是個難題,人們稱之為「C值謎」。
不過這些不含蛋白質編碼的脫氧核醣核酸序列,仍可能合成出具有功能的非編碼RNA分子,用以調控基因表現。
T7RNA聚合酶(藍色)以脫氧核醣核酸模板(橙色)為依據,合成mRNA(綠色)。
染色體中的某些非編碼脫氧核醣核酸序列,本身具有結構上的功能。
例如一般只帶有少量基因的端粒與著絲粒,對於染色體的穩定性及機能而言顯得相當重要。
人類體內有一類大量存在的非編碼脫氧核醣核酸,稱為偽基因,
是一些因突變累積而變得殘缺無用的基因複製品。
這些序列通常只可算是分子化石,不過有時候也會因為基因重複與趨異演化,而成為新基因裡的新遺傳物質。
轉錄與轉譯
基因是指一段含有遺傳訊息,且可影響生物體表現型的脫氧核醣核酸序列。
基因裡的脫氧核醣核酸鹼基序列決定了信使RNA的序列,而信使RNA的序列又決定了蛋白質的序列。
轉譯作用可依據基因所含有的核苷酸序列,以及遺傳密碼規則,生產出對應的蛋白質胺基酸序列。
遺傳密碼的組成單位稱為密碼子,是含有三個字母的「指令」,這些單位則由三個核苷酸組成,例如ACT、CAG或TTT。
在轉錄作用中,基因裡的密碼子會在RNA聚合酶的作用下,複製成為信使RNA。
之後核醣體會幫助帶著胺基酸的轉移RNA與信使RNA進行鹼基配對,進而將信使RNA解碼。
由於組成密碼子的鹼基共有四種,且以三字母為一單位,因此可能存在的密碼子一共有64種。
與這些密碼子對應的標準胺基酸有20種,因此大多數胺基酸對應了一種以上的密碼子。
另外有三個密碼子稱為「終止密碼子」或「無義密碼子」,是編碼區域的末端,
分別是TAA、TGA與TAG,這是屬於DNA上的終止密碼。
而在mRNA上的則是UAG
UAA UGA
當轉譯到達這三組密碼子時就會停止轉譯,並進行下一步的修飾
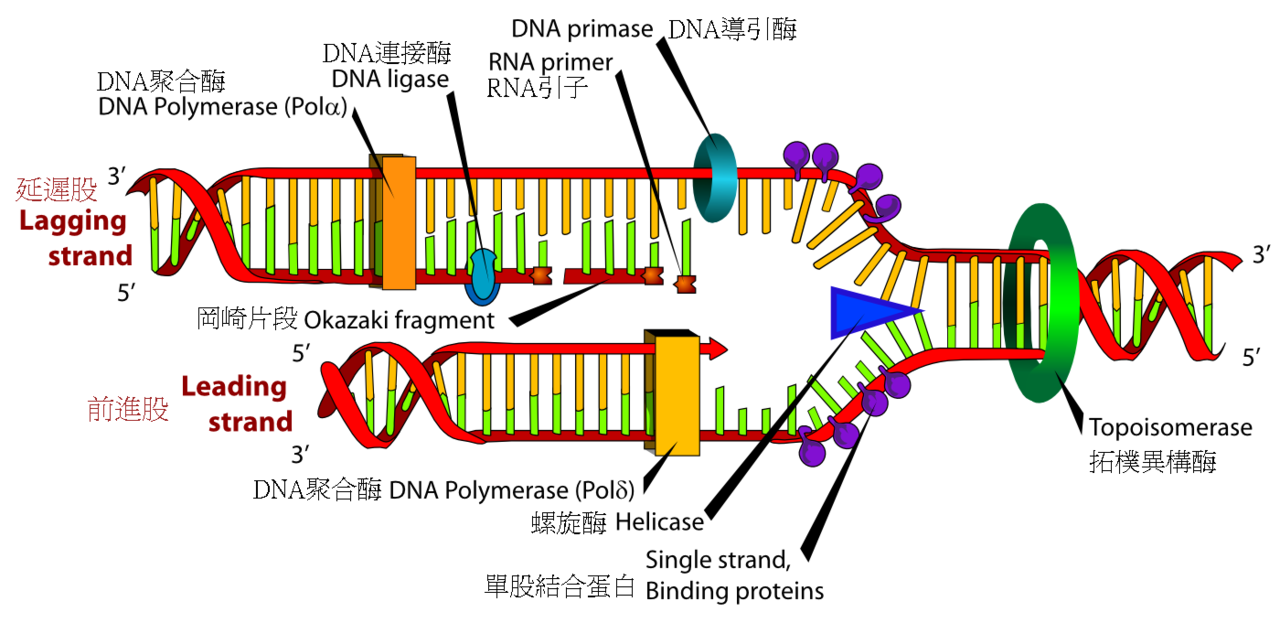
圖為脫氧核醣核酸複製,首先螺旋酶與拓撲異構酶將雙螺旋解開,接著一個DNA聚合酶負責合成前進股;
另一個則與延遲股結合,製造一些不連續的岡崎片段,再由脫氧核醣核酸連接酶將其黏合。
複製
生物個體成長需要經歷細胞分裂,當細胞進行分裂時,必須將自身基因組中的脫氧核醣核酸複製,
才能使子細胞擁有和親代相同的遺傳訊息。
脫氧核醣核酸的雙股結構可供脫氧核醣核酸複製機制進行,在此複製過程中, 兩條長鏈會先分離,
之後一種稱為DNA聚合酶的酵素,會分別以兩條長鏈為依據,合成出互補的脫氧核醣核酸序列。
酵素可找出正確的外來互補鹼基,並將其結合到模板長鏈上,進而製造出新的互補長鏈。
由於脫氧核醣核酸聚合酶只能以5'到3'的方向合成脫氧核醣核酸鏈,
因此雙螺旋中平行但方向相反的兩股,具有不同的合成機制。
舊長鏈上的鹼基序列決定了新長鏈上的鹼基序列,使細胞得以獲得完整的脫氧核醣核酸複製品。
與蛋白質的交互作用
脫氧核醣核酸若要發揮其功用,必須仰賴與蛋白質之間的交互作用,有些蛋白質的作用不具專一性,
有些則只專門與個別的脫氧核醣核酸序列結合。
聚合酶在各類酵素中尤其重要,此種蛋白質可與脫氧核醣核酸結合,並作用於轉錄或脫氧核醣核酸複製過程。
脫氧核醣核酸結合蛋白
結構蛋白可與脫氧核醣核酸結合,是非專一性脫氧核醣核酸-蛋白質交互作用的常見例子。
染色體中的結構蛋白與脫氧核醣核酸組合成複合物,使脫氧核醣核酸組織成緊密結實的染色質構造。
對真核生物來說,染色質是由脫氧核醣核酸與一種稱為組織蛋白的小型鹼性蛋白質所組合而成;
而原核生物體內的此種結構,則摻雜了多種類型的蛋白質。
雙股脫氧核醣核酸可在組織蛋白的表面上附著並纏繞整整兩圈,以形成一種稱為核小體的盤狀複合物。
組織蛋白裡的鹼性殘基,與脫氧核醣核酸上的酸性糖磷酸骨架之間可形成離子鍵,
使兩者發生非專一性交互作用,也使複合物中的鹼基序列相互分離。
在鹼性胺基酸殘基上所發生的化學修飾有甲基化、磷酸化與乙醯化等,
這些化學作用可使脫氧核醣核酸與組織蛋白之間的作用強度發生變化,
進而使脫氧核醣核酸與轉錄因子接觸的難易度改變,影響轉錄作用的速率。
其他位於染色體內的非專一性脫氧核醣核酸結合蛋白,還包括一種能優先與脫氧核醣核酸結合,
並使其扭曲的高移動性群蛋白。
這類蛋白質可以改變核小體的排列方式,產生更複雜的染色質結構。
脫氧核醣核酸結合蛋白中有一種專門與單股脫氧核醣核酸結合的類型,稱為單股脫氧核醣核酸結合蛋白。
人類的複製蛋白A是此類蛋白中獲得較多研究的成員,作用於多數與解開雙螺旋有關的過程,
包括脫氧核醣核酸複製、重組以及脫氧核醣核酸修復。
這類結合蛋白可固定單股脫氧核醣核酸,使其變得較為穩定,以避免形成莖環(stem-loop),或是因為核酸酶的作用而水解。
λ抑制子是一類具螺旋-轉角-螺旋結構的轉錄因子,可與脫氧核醣核酸目標結合。
相對而言,其他的蛋白質則只能與特定的脫氧核醣核酸序列進行專一性結合。
大多數關於此類蛋白質的研究集中於各種可調控轉錄作用的轉錄因子。
這類蛋白質中的每一種,都能與特定的脫氧核醣核酸序列結合,
進而活化或抑制位於啟動子附近序列的基因轉錄作用。
轉錄因子有兩種作用方式,第一種可以直接或經由其他中介蛋白質的作用,
而與負責轉錄的RNA聚合酶結合,再使聚合酶與啟動子結合,並開啟轉錄作用。
第二種則與專門修飾組織蛋白的酵素結合於啟動子上,使脫氧核醣核酸模板與聚合酶發生接觸的難度改變。
由於目標脫氧核醣核酸可能散佈在生物體中的整個基因組中,
因此改變一種轉錄因子的活性可能會影響許多基因的運作。
這些轉錄因子也因此經常成為信號傳遞過程中的作用目標,也就是作為細胞反映環境改變,
或是進行分化和發育時的媒介。
具專一性的轉錄因子會與脫氧核醣核酸發生交互作用,使脫氧核醣核酸鹼基的周圍產生許多接觸點,
讓其他蛋白質得以「讀取」這些脫氧核醣核酸序列。
多數的鹼基交互作用發生在大凹槽,也就是最容易從外界接觸鹼基的部位。
遺傳重組
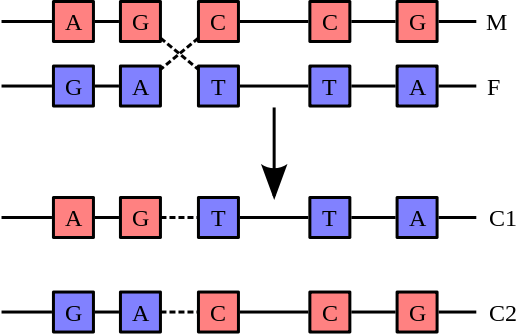
重組過程中,兩條染色體(M與F)斷裂之後又重新接合,產生兩條重新排列過的染色體(C1與C2)。
各條脫氧核醣核酸螺旋間的交互作用不常發生,在人類細胞核裡的每個染色體,
各自擁有一塊稱作「染色體領域」的區域。
染色體之間在物理上的分離,對於維持脫氧核醣核酸資訊儲藏功能的穩定性而言相當重要。
不過染色體之間有時也會發生重組,在重組的過程中,會進行染色體互換:
首先兩條脫氧核醣核酸螺旋會先斷裂,之後交換其片段,最後再重新黏合。
重組作用使染色體得以互相交換遺傳訊息,並產生新的基因組合,進而增加自然選擇的效果,
且可能對蛋白質的演化產生重要影響。
遺傳重組也參與脫氧核醣核酸修復作用,尤其是當細胞中的脫氧核醣核酸發生斷裂的時候。
同源重組是最常見的染色體互換方式,可發生於兩條序列相類似的染色體上。
而非同源重組則對細胞具有傷害性,會造成染色體易位與遺傳異常。
可催化重組反應的酵素,如RAD51,稱為「重組酶」。
重組作用的第一個步驟,是內切酶作用,或是脫氧核醣核酸的損壞所造成的脫氧核醣核酸雙股斷裂。
重組酶可催化一系列步驟,使兩條螺旋結合產生Holliday交叉。
其中每條螺旋中的單股脫氧核醣核酸,皆與另一條螺旋上與之互補的脫氧核醣核酸連結在一起,
進而形成一種可於染色體內移動的交叉形構造,造成脫氧核醣核酸鏈的互換。
重組反應最後會因為交叉結構的斷裂,以及脫氧核醣核酸的重新黏合而停止。
脫氧核醣核酸生物代謝的演化
脫氧核醣核酸所包含的遺傳訊息,是所有現代生命機能,以及生物生長與繁殖的基礎。
不過目前尚未明瞭在長達40億年的生命史中,脫氧核醣核酸究竟是何時出現並開始發生作用。
有一些科學家認為,早期的生命形態有可能是以RNA作為遺傳物質。
RNA可能在早期細胞代謝中扮演主要角色,一方面可傳遞遺傳訊息;另一方面也可作為核醣酶的一部分,進行催化作用。
在古代RNA世界裡,核酸同時具有催化與遺傳上的功能,而這些分子後來可能演化成為目前以四種核苷酸組成遺傳密碼的形式,
這是因為當鹼基種類較少時,複製的精確性會增加;而鹼基種類較多時,增加的則是核酸的催化效能。
兩種可達成不同目的功能最後在四種鹼基的情形下達到最合適數量。
不過關於這種古代遺傳系統並沒有直接證據,且由於脫氧核醣核酸在環境中無法存留超過一百萬年,
在溶液中又會逐漸降解成短小的片段,因此大多數化石中並無脫氧核醣核酸可供研究。
即使如此,仍有一些聲稱表示已經獲得更古老的DNA,其中一項研究表示,
已從存活於2億5千萬年古老的鹽類晶體中的細菌分離出脫氧核醣核酸,但此宣布引起了討論與爭議。(
http://zh.wikipedia.org/wiki/%E8%84%B1%E6%B0%A7%E6%A0%B8%E7%B3%96%E6%A0%B8%E9%85%B8